AG Cytoskeletal Dynamics
Fast alle Vorgänge in mehrzelligen Organismen werden durch ein –im Optimalfall perfektes- Zusammenspiel zwischen einzelnen Zellen bestimmt. Die Kommunikation zwischen Zellen (Signaltransduktion) kann zu sehr dynamischen Prozessen wie Zellteilung, Endo- und Exocytose sowie Bewegung (Migration und Invasion) von Zellen führen. Zu den beweglichsten Zellen des menschlichen Organismus zählen die Immunzellen, die durch Chemokine an den Ort der Infektion gelockt werden. Aber auch Nervenzellen (Neurone), Knochenzellen (Osteoblasten, Osteoklasten), Haut- und Bindegewebszellen sind sehr beweglich. Während der Migration erfolgt eine steter Auf- und Abbau des Zytoskeletts, das aus Intermediär- und Aktinfilamenten sowie aus Mikrotubuli besteht. Die treibende Kraft für die Migration wird durch die Dynamik der Aktinfilamente bereitgestellt. Hierbei werden am vorderen Ende der Zellen Aktinfilamente auf- und am hinteren Ende abgebaut. Diese Dynamik der Aktinfilamente wird durch Aktinbindeproteine kontrolliert. Mikrotubuli dagegen regulieren hauptsächlich die Bewegung von Spindelfasern und den Transport von Vesikeln. Deshalb sind die Zellteilung und die Proteinsekretion zum Großteil durch Mikrotubuli gesteuert. Ähnlich wie bei Aktinfilamenten, wird die Mikrotubuli-Dynamik durch assoziierte Proteine reguliert.
Ist die Regulation und/oder Bindung von Zytoskelett-assoziierten Proteinen gestört, kann es zu Störungen der Zellphysiologie kommen. Dies ist besonders bei Autoimmunkrankheiten, neuronalen Störungen und bei metastasierenden Tumorzellen von großer Bedeutung. Aus diesem Grund ist es ein sehr interessanter Ansatz nach Zytoskelett-assoziierten Proteinen zu suchen, die essentiell für die Funktion von Immunzellen, Neuronen und Tumorzellen sind. Je nach Krankheitsbild sollte eine Stimulation oder Hemmung dieser Proteine die Störung abschwächen.
Unsere Gruppe untersucht hauptsächlich die Funktion von Zytoskelett-assoziierten Proteinen bei der Metastasierung von Tumorzellen. Es gelang uns bereits Proteine zu identifizieren, die an der Regulation dieser Prozesse beteiligt sind. Unter diesen Proteinen wurde die Inositol-1,4,5-trisphosphat 3-Kinase-A (ITPKA) besonders gut untersucht. Darüber hinaus interessieren wir uns generell dafür wie das Zytoskelett von Tumorzellen reguliert wird. Ein genaues Verständnis darüber ermöglicht es, die Zytoskelett-Dynamik in Tumorzellen zu hemmen und damit die Metastasierung von Tumorzellen zu blockieren.
Im Folgenden sind kurz unsere aktuellen Projekte zusammen gefasst:
Inositol-1,4,5-trisphosphat 3-Kinase-A (ITPKA)
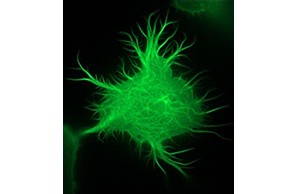
Wir forschen bereits seit mehr als zehn Jahren an der Rolle der physiologisch nur in Neuronen exprimierten Inositol-1,4,5-trisphosphat 3-Kinase-A (InsP3Kinase oder ITPKA) bei der Tumorgenese. Umfangreiche Untersuchungen ergaben, dass ITPKA in Lungenkarzinomzellen überexprimiert ist und die Metastasierung dieser Zellen kontrolliert (Windhorst et al., 2008; 2010; 2011, 2017, Küster et al., 2023). Dies geschieht durch die InsP3Kinase-vermittelte Regulation von zytosolischen Calciumsignalen sowie durch die Vernetzung von Aktinfilamenten (s. Bild rechts und Ashour et al., 2015).
Die Überexpression von ITPKA in Lungenkarzinomzellen wird durch Fehlregulation des Repressor-Element-1-Silencing-Transcription-Factor REST/NRSF induziert (Chang et al., 2011). Dieser Transkriptionsrepressor führt normalerweise zur Repression von nicht-neuronalen Genen in nicht-neuronalem Gewebe. In vielen Tumorzellen ist REST/NRSF fehlreguliert, so dass es zur Expression von neuronalen Genen kommt. Tumorzellen nutzen neuronale Proteine u.a. zur autokrinen Stimulation und verbesserten Migration und Invasion.
Es gelang uns bereits Hemmstoffe gegen die InsP3Kinase-Aktivität zu identifizieren, von denen BIP-4 (Schröder et al., 2013; 2015; Paraschiakos et al., 2021) als Leitstruktur verwendet werden kann, um Medikamente zu entwickeln, die eine Hemmung der InsP3Kinase-Aktivität in Tumorzellen bewirken.
Zurzeit arbeiten wir daran, Hemmstoffe gegen die Aktin-Crosslinking-Aktivität von ITPKA zu suchen. Hierzu verfolgen wir zwei Ansätze (1) die Selektion von Sybodies (synthetische Nanobodies) gegen die Aktinbinde-Domäne von ITPKA, (2) die Identifizierung von „small molecules“ oder Peptidomimetics, die spezifisch die Interaktion zwischen ITPKA und Aktin blockieren.
Gibt es Master-Aktinbindeproteine, die essentiell für die Metastasierung von Tumorzellen sind?
Die meisten Tumorzell-Populationen sind heterogen und plastisch. Aus diesem Grund ist es schwierig, ein einziges Protein zu identifizieren, das essentiell für die Metastasierung aller Tumorzellen in einer Population ist. Außerdem verändern sich Tumorzellen während der Metastasierung dramatisch (Schäfer et al., 2023). In der folgenden Abbildung ist diese Plastizität dargestellt:
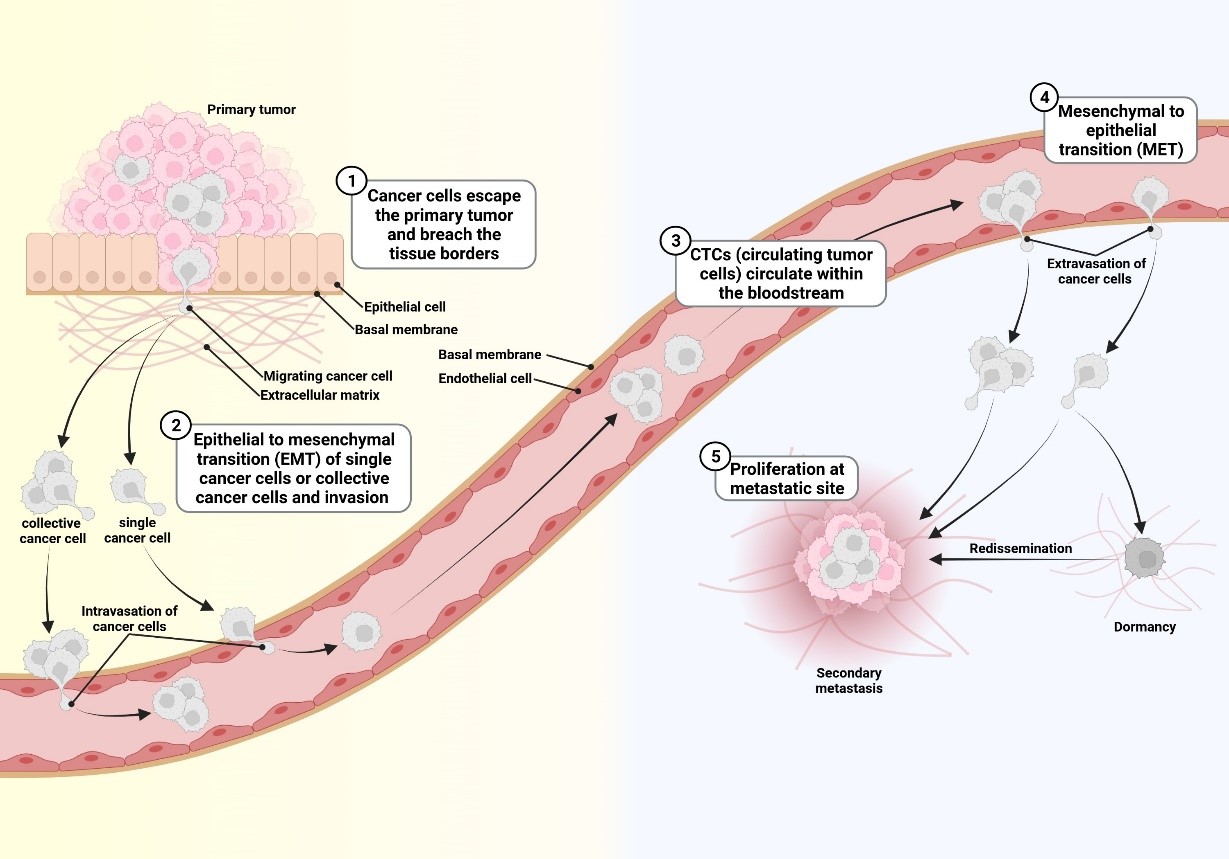
Diese Tatsache erschwert die zielgerichtete Therapie von Tumorzellen durch Hemmung nur eines Proteins (z.B Her2 in Mammakarzinomzellen) und deshalb werden Krebspatient:innen in den meisten Fällen immer noch mit einer Kombination aus zielgerichteter- und Chemotherapie sowie durch Bestrahlung behandelt. Neuerdings werden auch Immuntherapien angeboten.
Da Tumorzellen während der Metastasierung darauf angewiesen sind, ihr Zytoskelett kontinuierlich auf- und abzubauen, benötigen sie unterschiedliche Aktinbindeproteine, die diesen Prozess vermitteln. Um zu prüfen, welches Aktinbindeprotein am stärksten in metastasierten Tumorzellen exprimiert ist, untersuchten wir in Kooperation mit der Gynäkologie und der Bioinformatik des UKE die mRNA Expression von Aktinbindeproteinen in Proben von Brustkrebspatientinnen. Hierbei stellte sich heraus, dass nur wenige Aktinbindeproteine signifikant in metastasierten Tumoren herauf reguliert sind. Die stärkste Korrelation zeigte das Aktinschneide-Protein Cofilin-1. Dieses Protein ist essentiell, um die Aktin-Dynamik in sich bewegenden Zellen aufrecht zu erhalten und spielt deshalb eine wichtige Rolle bei der Metastasierung.
Zurzeit analysieren wir Patientenproben per Western-Blotting um zu prüfen, ob Cofilin-1 wirklich in allen Proben von metastasierten Tumoren exprimiert ist. Sollte dies tatsächlich der Fall sein, werden wir im nächsten Schritt prüfen, ob die Cofilin-Expression auch während der Metastasierung konstant bleibt, oder sich in plastischen Zellen verändert. Falls wir finden sollten, dass Cofilin-1 tatsächlich von allen Zellen in einer Tumorzellpopulation und auch in allen Tumortypen exprimiert ist, wäre Cofilin-1 ein Master-Aktinbindeprotein und sein Hemmung sollte folglich die Metastasierung hemmen. Wahrscheinlicher ist allerding, dass Tumorzellen unterschiedliche Strategien nutzen, um zu metastasieren und deshalb auch unterschiedlicher Aktinbindeproteine exprimieren.
Die fehlerhafte Expression von Mikrotubuli-assoziierten Proteinen steigert die chromosomale Instabilität von Tumorzellen
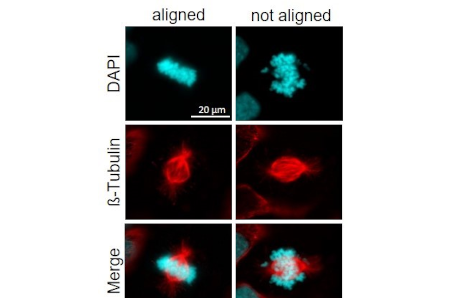
Chromosomale Instabilität ist ein zellulärer Prozess bei dem entweder ganze Chromosomen oder Teile davon nach der Zellteilung ungleichmäßig auf die Tochterzellen aufgeteilt werden. Das kann zum „gain of function“ oder „loss of function“ führen, so das Tumorzellen z.B. Gene dazu gewinnen können, die für Onkogene codieren oder Tumorsuppressor-codierende Gene verlieren. Nach Selektionsdruck überleben also nur solche Tumorzellen, die ein optimal angepasstes Genom haben. Diese starke genomische Plastizität erlaubt es den Zellpopulationen sich immer wieder an neue Gegebenheiten anzupassen, was die Tumortherapie ungemein erschwert. Ein wichtiger Faktor, der zur chromosomalen Instabilität führen kann, ist eine fehlerhafte Regulation der Spindel-Mikrotubuli-Dynamik. Die mitotischen Spindelfasern binden die Chromosomen in der Metaphase und ordnen sie in der Äquatorial-Ebene an („aligned“, Bild rechts).
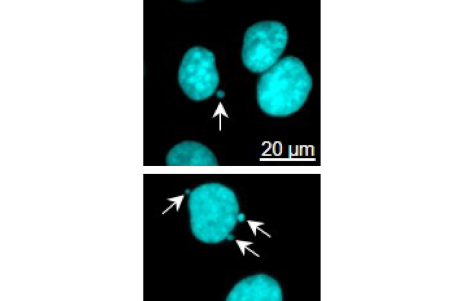
Danach ziehen die Mikrotubuli die Chromosomen zu den Zellpolen und gewährleisten so, dass die Chromosomen gleichmäßig auf die Tochterzellen verteilt werden. Funktieren die Spindel-Mikrotubuli nicht richtig kann es passieren, dass die Chromosomen nicht korrekt angeordnet sind („not aligned“) und nachfolgend in der Anaphase nicht gleichmäßig zu den Zellpolen gezogen werden. Aus diesem Grund werden die Chromosomen ungleich zwischen den Tochterzellen aufgeteilt oder verbleiben im Zytosol. Die zytosolischen Chromosomen werden von einer Kernmembran umschlossen und etablieren sich als „Mikronuclei“ (s. Pfeile in Abb. rechts). Diese Mikronuclei können brechen, so dass sich DNA im Zytosol anreichert und eine Immunreaktion hervorruft. Die zytosolische DNA wird von der cyclic GMP-AMP synthase (cGAS) erkannt und zyklisches GMP-AMP aktiviert nachfolgend den Stimulator of INterferon Genes (STING). Schließlich sezernieren Zellen mit hohen zytosolischen DNA Konzentrationen Interferone und stimulieren auf diese Weise vor allem T- und dendritische Zellen.
Aus diesem Grund ist es wahrscheinlich, dass chromosomal instabile Zellen zu einer Stimulation von Immunzellen führen und deshalb möglicherweise besser auf Immunzell-Therapien reagieren als chromosomal stabile Zellen. Zurzeit sind wir dabei diese Hypothese zu prüfen.
Regulation der Exosomen-Biogenese durch Mikrotubuli-assoziierte Proteine
Exosomen sind kleine Vesikel, die von Zellen sezerniert werden, um Informationen an Nachbarzellen weiterzugeben. Im Fall von Tumorzellen kann die Tumormikroumgebung durch Exosomen so vorbereitet werden, dass sich Tumorzellen in fremden Geweben besser ausbreiten können. Zurzeit wird angenommen, dass die Exosomen-Biogenese mit der Einschnürung der Endosomen-Membran beginnt. Die Endosomen reifen dann zu „Multivesicular-Bodies“ (MVB), die intraluminale Vesikel (ILV) enthalten. Bestimmte MVB-Populationen verschmelzen schließlich mit der Plasmamembran und entlassen die ILV als Exosomen ins Medium. Interessanterweise wird bei der Bildung der ILV Material aus dem Zytoplasma (Protein, Nukleinsäuren und Metabolite) eingeschleust und über die Exosomen an Nachbarzellen abgegeben.
Wir konnten kürzlich zeigen, dass eine bestimmte posttranslationale Modifikation von Mikrotubuli dazu führt, dass einige MVB-Populationen schneller innerhalb der Zellen transportiert und entsprechend mehr Exosomen sezerniert wurden. Interessanterweise manipulierten diese Tumorzell-Exosomen Zellen der Blut-Hirn-Schranke derartig, dass sie durchlässiger wurde und Tumorzellen besser an ihnen haften konnten (Arnold et al., 2021).
Zurzeit untersuchen wir die Rolle eines weiteren Mikrotubuli-assoziierten Proteins bei der Biogenese von Exosomen. Dieses Protein scheint die Aktivität von ESCRT (endosomal sorting complexes required for transport)-Proteinen in Tumorzellen zu stimulieren und damit die ILV-Synthese zu beschleunigen. Da das Protein nicht in normalen Zellen exprimiert wird, scheint es ein Tumorzell-spezifischer Promotor der Exosomen-Synthese zu sein. Zurzeit sind wir dabei diese Annahme zu überprüfen.
Zusammengefasst zeigen unsere Projekte deutlich, das Zytoskelett-assoziierte Proteine essentiell für unterschiedliche zelluläre Prozesse sind. Folglich gibt es vielfältige Möglichkeiten durch Hemmung solcher Proteine die Metastasierung von Tumorzellen zu hemmen.
Techniken/Methoden
- Klonierungen und Proteinreinigungen
- Extraktion von DNA, mRNA und Proteinen
- Enzymkinetische Assays
- Bestimmung der cytosolischen Calciumkonzentration
- Westernblotting, Immunpräzipitation, Immuncytologie und Histololgie
- Migrations- Invasions- und Adhäsions-Assays
- Transiente und stabile Überexpression sowie shRNA-vermittelte Herunterregulation von Proteinen.
- Diverse Techniken zur Analyse der Aktin- und Mikrotubuli Dynamik in zellfreien Systemen
- Fluoreszenzmikroskopie
Ausbildung
Betreuung von Bachelor-, Master- und Doktorarbeiten.
Publikationen seit 2002
2023
Cryo-EM structures of actin binding proteins as tool for drug discovery
Dahlstroem C, Paraschiakos T, Sun H, Windhorst S
BIOCHEM PHARMACOL. 2023;214:115680.
The actin bundling activity of ITPKA mainly accounts for its migration-promoting effect in lung cancer cells
Küster L, Paraschiakos T, Karakurt K, Schumacher U, Diercks B, Windhorst S
BIOSCIENCE REP. 2023;43(2):.
Ex Vivo Model of Neuroblastoma Plasticity
Schäfer P, Muhs S, Turnbull L, Garwal P, Maar H, Yorgan T, Tolosa E, Lange T, Windhorst S
CANCERS. 2023;15(4):.
Structural basis for subversion of host cell actin cytoskeleton during Salmonella infection
Yuan B, Scholz J, Wald J, Thuenauer R, Hennell James R, Ellenberg I, Windhorst S, Faix J, Marlovits T
SCI ADV. 2023;9(49):eadj5777.
2022
DIAPH1 facilitates paclitaxel-mediated cytotoxicity of ovarian cancer cells
Flat W, Borowski S, Paraschiakos T, Blechner C, Windhorst S
BIOCHEM PHARMACOL. 2022;197:.
Key Role of Hyaluronan Metabolism for the Development of Brain Metastases in Triple-Negative Breast Cancer
Hamester F, Stürken C, Legler K, Eylmann K, Möller K, Roßberg M, Gorzelanny C, Bauer A, Windhorst S, Schmalfeldt B, Laakmann E, Müller V, Witzel I, Oliveira-Ferrer L
CELLS-BASEL. 2022;11(20):.
2021
2-Methoxyestradiol and its derivatives inhibit store-operated Ca2+ entry in T cells: identification of a new and potent inhibitor
Löhndorf A, Hosang L, Dohle W, Odoardi F, Waschkowski S, Rosche A, Bauche A, Winzer R, Tolosa E, Windhorst S, Marry S, Flügel A, Potter B, Diercks B, Guse A
BBA-MOL CELL RES. 2021;1868(6):118988.
DIAPH1 regulates chromosomal instability of cancer cells by controlling microtubule dynamics
Miao S, Schäfer P, Nojszewski J, Meyer F, Windhorst S
EUR J CELL BIOL. 2021;100(3):.
Overexpression of Lin28A in neural progenitor cells in vivo does not lead to brain tumor formation but results in reduced spine density
Middelkamp M, Ruck L, Krisp C, Sumisławski P, Mohammadi B, Dottermusch M, Meister V, Küster L, Schlüter H, Windhorst S, Neumann J
ACTA NEUROPATHOL COM. 2021;9(1):185.
Mechanism of BIP-4 mediated inhibition of InsP3Kinase-A
Paraschiakos T, Flat W, Chen Y, Kirchmair J, Windhorst S
BIOSCIENCE REP. 2021;41(7):.
Oncogenic activity and cellular functionality of melanoma associated antigen A3
Schäfer P, Paraschiakos T, Windhorst S
BIOCHEM PHARMACOL. 2021;192:.
2020
Tubulin Tyrosine Ligase Like 4 (TTLL4) overexpression in breast cancer cells is associated with brain metastasis and alters exosome biogenesis
Arnold J, Schattschneider J, Blechner C, Krisp C, Schlüter H, Schweizer M, Nalaskowski M, Oliveira-Ferrer L, Windhorst S
J EXP CLIN CANC RES. 2020;39(1):205.
Physiological relevance of the neuronal isoform of inositol-1,4,5-trisphosphate 3-kinases in mice
Blechner C, Becker L, Fuchs H, Rathkolb B, Prehn C, Adler T, Calzada-Wack J, Garrett L, Gailus-Durner V, Morellini F, Conrad S, Hölter S, Wolf E, Klopstock T, Adamski J, Busch D, de Angelis M, Schmeisser M, Windhorst S
NEUROSCI LETT. 2020;735:135206.
Conserved Tao Kinase Activity Regulates Dendritic Arborization, Cytoskeletal Dynamics, and Sensory Function in Drosophila
Hu C, Kanellopoulos A, Richter M, Petersen M, Konietzny A, Tenedini F, Hoyer N, Cheng L, Poon C, Harvey K, Windhorst S, Parrish J, Mikhaylova M, Bagni C, Calderon de Anda F, Soba P
J NEUROSCI. 2020;40(9):1819-1833.
Modeling Spontaneous Bone Metastasis Formation of Solid Human Tumor Xenografts in Mice
Labitzky V, Baranowsky A, Maar H, Hanika S, Starzonek S, Ahlers A, Stübke K, Koziolek E, Heine M, Schäfer P, Windhorst S, Jücker M, Riecken K, Amling M, Schinke T, Schumacher U, Valentiner U, Lange T
CANCERS. 2020;12(2):.
Characterization of the substrate specificity of the inositol 5-phosphatase SHIP1
Nelson N, Wundenberg T, Lin H, Rehbach C, Horn S, Windhorst S, Jücker M
BIOCHEM BIOPH RES CO. 2020;524(2):366-370.
Mice lacking plastin-3 display a specific defect of cortical bone acquisition
Yorgan T, Sari H, Rolvien T, Windhorst S, Failla A, Kornak U, Oheim R, Amling M, Schinke T
BONE. 2020;130:115062.
2019
Inositol hexakisphosphate increases the size of platelet aggregates
Brehm M, Klemm U, Rehbach C, Erdmann N, Kolšek K, Lin H, Aponte-Santamaría C, Gräter F, Rauch B, Riley A, Mayr G, Potter B, Windhorst S
BIOCHEM PHARMACOL. 2019;161:14-25.
New options of cancer treatment employing InsP6
Brehm M, Windhorst S
BIOCHEM PHARMACOL. 2019;163:206-214.
The formin Drosophila homologue of Diaphanous2 (Diaph2) controls microtubule dynamics in colorectal cancer cells independent of its FH2-domain
Grueb S, Muhs S, Popp Y, Schmitt S, Geyer M, Lin Y, Windhorst S
SCI REP-UK. 2019;9(1):5352.
Radial somatic F-actin organization affects growth cone dynamics during early neuronal development
Meka D, Scharrenberg R, Zhao B, Kobler O, König T, Schaefer I, Schwanke B, Klykov S, Richter M, Eggert D, Windhorst S, Dotti C, Kreutz M, Mikhaylova M, Calderon de Anda F
EMBO REP. 2019;20(12):e47743.
The Actin Binding Protein Plastin-3 Is Involved in the Pathogenesis of Acute Myeloid Leukemia
Velthaus A, Cornils K, Hennigs J, Grüb S, Stamm H, Wicklein D, Bokemeyer C, Heuser M, Windhorst S, Fiedler W, Wellbrock J
CANCERS. 2019;11(11):.
2018
Effect of the actin- and calcium-regulating activities of ITPKB on the metastatic potential of lung cancer cells
Bäder S, Glaubke E, Grüb S, Muhs S, Wellbrock J, Nalaskowski M, Lange T, Windhorst S
BIOCHEM J. 2018;475(12):2057-2071.
Differential Proteome Analysis of Human Neuroblastoma Xenograft Primary Tumors and Matched Spontaneous Distant Metastases
Hänel L, Gosau T, Maar H, Valentiner U, Schumacher U, Riecken K, Windhorst S, Hansen N, Heikaus L, Wurlitzer M, Nolte I, Schlüter H, Lange T
SCI REP-UK. 2018;8(1):13986.
Clinical relevance of cytoskeleton associated proteins for ovarian cancer
Schiewek J, Schumacher U, Lange T, Joosse S, Wikman H, Pantel K, Mikhaylova M, Kneussel M, Linder S, Schmalfeldt B, Oliveira-Ferrer L, Windhorst S
J CANCER RES CLIN. 2018;144(11):2195-2205.
2017
Strong fascin expression promotes metastasis independent of its F-actin bundling activity
Heinz L, Muhs S, Schiewek J, Grüb S, Nalaskowski M, Lin Y, Wikman H, Oliveira-Ferrer L, Lange T, Wellbrock J, Konietzny A, Mikhaylova M, Windhorst S
ONCOTARGET. 2017;8(66):110077-110091.
Tight Junction Proteins Claudin-1 and Occludin Are Important for Cutaneous Wound Healing
Volksdorf T, Heilmann J, Eming S, Schawjinski K, Zorn-Kruppa M, Ueck C, Vidal-Y-Sy S, Windhorst S, Jücker M, Moll I, Brandner J
AM J PATHOL. 2017;187(6):1301-1312.
Inositol-1,4,5-trisphosphate 3-kinase-A (ITPKA) is frequently over-expressed and functions as an oncogene in several tumor types
Windhorst S, Song K, Gazdar A
BIOCHEM PHARMACOL. 2017;137:1-9.
Microtubules Modulate F-actin Dynamics during Neuronal Polarization
Zhao B, Meka P, Scharrenberg R, König T, Schwanke B, Kobler O, Windhorst S, Kreutz M, Mikhaylova M, Calderon de Anda F
SCI REP-UK. 2017;7(1):9583.
2016
Smooth Muscle-Alpha Actin Inhibits Vascular Smooth Muscle Cell Proliferation and Migration by Inhibiting Rac1 Activity
Chen L, DeWispelaere A, Dastvan F, Osborne W, Blechner C, Windhorst S, Daum G
PLOS ONE. 2016;11(5):e0155726.
Control of aromatase in hippocampal neurons
Fester L, Brandt N, Windhorst S, Pröls F, Bläute C, Rune G
J STEROID BIOCHEM. 2016;160:9-14.
Inositol-1,4,5-trisphosphate-3-kinase-A controls morphology of hippocampal dendritic spines
Köster J, Leggewie B, Blechner C, Brandt N, Fester L, Rune G, Schweizer M, Kindler S, Windhorst S
CELL SIGNAL. 2016;28(1):83-90.
Diaphanous-related formin 1 as a target for tumor therapy
Lin Y, Windhorst S
BIOCHEM SOC T. 2016;44(5):1289-1293.
2015
The catalytic domain of inositol-1,4,5-trisphosphate 3-kinase-a contributes to ITPKA-induced modulation of F-actin
Ashour D, Pelka B, Jaaks P, Wundenberg T, Blechner C, Zobiak B, Failla A, Windhorst S
CYTOSKELETON. 2015;72(2):93-100.
Drosophila homologue of Diaphanous 1 (DIAPH1) controls the metastatic potential of colon cancer cells by regulating microtubule-dependent adhesion
Lin Y, Bhuwania R, Gromova K, Failla A, Lange T, Riecken K, Linder S, Kneussel M, Izbicki J, Windhorst S
ONCOTARGET. 2015;6(21):18577-89.
Ex vivo aorta patch model for analysis of cellular adhesion
Lin Y, Thata R, Failla A, Geissen M, Daum G, Windhorst S
TISSUE CELL. 2015;47(3):266-272.
The new InsP3Kinase inhibitor BIP-4 is competitive to InsP3 and blocks proliferation and adhesion of lung cancer cells
Schröder D, Tödter K, Gonzalez B, Franco-Echevarría E, Rohaly G, Blechner C, Lin H, Mayr G, Windhorst S
BIOCHEM PHARMACOL. 2015;96(2):143-50.
2014
Expression of DIAPH1 is up-regulated in colorectal cancer and its down-regulation strongly reduces the metastatic capacity of colon carcinoma cells
Lin Y, Izbicki J, König A, Habermann J, Blechner C, Lange T, Schumacher U, Windhorst S
INT J CANCER. 2014;134(7):1571-82.
Expression, Purification and preliminary X-Ray diffraction of a pathogenic bacterial protease from Stenotrophomonas maltophilia
Negm A, Windhorst S, Betzel C, Akrem A, Weber W
World J Pharm Pharm Sci. 2014;3(10):13-23.
Cellular internalisation of an inositol phosphate visualised by using fluorescent InsP5
Riley A, Windhorst S, Lin H, Potter B
CHEMBIOCHEM. 2014;15(1):57-67.
2013
Expression Regulation of the Metastasis-Promoting Protein InsP3-Kinase-A in Tumor Cells.
Chang L, Schwarzenbach H, Meyer-Staeckling S, Brandt B, Mayr G, Weitzel J, Windhorst S
MOL CANCER RES. 2013.
Combined targeting of AKT and mTOR using MK-2206 and RAD001 is synergistic in the treatment of cholangiocarcinoma
Ewald F, Grabinski N, Grottke A, Windhorst S, Nörz D, Carstensen L, Staufer K, Hofmann B, Diehl F, David K, Schumacher U, Nashan B, Jücker M
INT J CANCER. 2013;133(9):2065-76.
Malignant H1299 tumour cells preferentially internalize iron-bound inositol hexakisphosphate
Helmis C, Blechner C, Lin H, Schweizer M, Mayr G, Nielsen P, Windhorst S
BIOSCIENCE REP. 2013;33(5):.
Identification of a new membrane-permeable inhibitor against inositol-1,4,5-trisphosphate-3-kinase A
Schröder D, Rehbach C, Seyffarth C, Neuenschwander M, Kries J, Windhorst S
BIOCHEM BIOPH RES CO. 2013;439(2):228-34.
Tumour cells can employ extracellular Ins(1,2,3,4,5,6)P(6) and multiple inositol-polyphosphate phosphatase 1 (MINPP1) dephosphorylation to improve their proliferation
Windhorst S, Lin H, Blechner C, Fanick W, Brandt L, Brehm M, Mayr G
BIOCHEM J. 2013;450(1):115-25.
2012
Inositol-1,4,5-trisphosphate 3-kinase A regulates dendritic morphology and shapes synaptic Ca2+ transients.
Windhorst S, Minge D, Bähring R, Hüser S, Schob C, Blechner C, Lin H, Mayr G, Kindler S
CELL SIGNAL. 2012;24(3):750-757.
2011
Human inositol 1,4,5-trisphosphate 3-kinase isoform B (IP3KB) is a nucleocytoplasmic shuttling protein specifically enriched at cortical actin filaments and at invaginations of the nuclear envelope.
Nalaskowski M, Fliegert R, Ernst O, Brehm M, Fanick W, Windhorst S, Lin H, Giehler S, Hein J, Lin Y, Mayr G
J BIOL CHEM. 2011;286(6):4500-4510.
Functional role of inositol-1,4,5-trisphosphate-3-kinase-A for motility of malignant transformed cells.
Windhorst S, Kalinina T, Schmid K, Blechner C, Kriebitzsch N, Hinsch R, Chang L, Herich L, Schumacher U, Mayr G
INT J CANCER. 2011;129(6):1300-1309.
2010
Inositol-1,4,5-trisphosphate-3-kinase-A is a new cell motility-promoting protein that increases the metastatic potential of tumour cells by two functional activities.
Windhorst S, Fliegert R, Blechner C, Möllmann K, Hosseini Z, Guenther T, Eiben M, Chang L, Lin H, Fanick W, Schumacher U, Brandt B, Mayr G
J BIOL CHEM. 2010;285(8):5541-5554.
2008
Ins(1,4,5)P3 3-kinase-A overexpression induces cytoskeletal reorganization via a kinase-independent mechanism.
Windhorst S, Blechner C, Lin H, Elling C, Nalaskowski M, Kirchberger T, Guse A, Mayr G
BIOCHEM J. 2008;414(3):407-417.
2007
Intracellular localization of human Ins(1,3,4,5,6)P5 2-kinase.
Brehm M, Schenk T, Zhou X, Fanick W, Lin H, Windhorst S, Nalaskowski M, Kobras M, Shears S, Mayr G
BIOCHEM J. 2007;408(3):335-345.
Radiosensitization of tumour cell lines by the polyphenol Gossypol results from depressed double-strand break repair and not from enhanced apoptosis.
Kasten-Pisula U, Windhorst S, Dahm-Daphi J, Mayr G, Dikomey E
RADIOTHER ONCOL. 2007;83(3):296-303.
2006
Subcellular localisation of human inositol 1,4,5-trisphosphate 3-kinase C: species-specific use of alternative export sites for nucleo-cytoplasmic shuttling indicates divergent roles of the catalytic and N-terminal domains.
Nalaskowski M, Windhorst S, Stockebrand M, Mayr G
BIOL CHEM. 2006;387(5):583-593.
2005
Antiproliferative plant and synthetic polyphenolics are specific inhibitors of vertebrate inositol-1,4,5-trisphosphate 3-kinases and inositol polyphosphate multikinase.
Mayr G, Windhorst S, Hillemeier K
J BIOL CHEM. 2005;280(14):13229-13240.
Letzte Aktualisierung aus dem FIS: 26.07.2024 - 00:33 Uhr